Piramidal hüceyrə

Piramidal hüceyrə və ya piramidal neyronlar — beyin qabığı, hipokampus və amigdala[1] da daxil olmaqla beynin bölgələrində tapılan çoxqütblü neyronların bir növü. Piramidal neyronlar məməlilərin beynindəki əsas həyəcanverici neyronlardır. Balıqlarda, quşlarda, sürünənlərdə də tapılır.
Piramidal neyronlar həm də ölümlə nəticələnən quduzluq infeksiyasında xarakterik əlamət olan Neqri cisimlərinin tapıldığı 2 hüceyrə tipindən biridir.[2]
Piramidal neyronlar ilk dəfə Santiaqo Ramon i Kajal tərəfindən kəşf edilmiş və tədqiq edilmişdir.[3][4] O vaxtdan bəri, piramidal neyronlar üzərində aparılan tədqiqatlar neyroplastiklikdən tutmuş koqnitiv funksiyaya qədər müxtəlif mövzulara yönəlmişdir.
Strukturu[redaktə | mənbəni redaktə et]
-

Yaşıl flüoresan zülal (yfz) ilə vizuallaşdırılan piramidal neyron
-
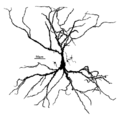
Hipokampusun piramidal hüceyrəsi


Piramidal neyronun əsas struktur xüsusiyyətlərindən biri konus formalı soma (piramidal hüceyrə cisimləri) və ya hüceyrə gövdəsidir ki, neyron onun adını daşıyır. Piramidal hüceyrənin digər əsas struktur xüsusiyyətləri tək akson, böyük apikal dendrit, çoxsaylı bazal dendritlər və dendritik tikanların olmasıdır.[5]
Apikal dendrit[redaktə | mənbəni redaktə et]
Apikal dendrit piramidal hüceyrənin somasının (perikarionun) yuxarı hissəsindən qalxır. Apikal dendrit tək, uzun və qalın dendritdir, somadan məsafə artdıqca bir neçə dəfə budaqlanır və qabıq səthinə doğru uzanır.
Bazal dendrit[redaktə | mənbəni redaktə et]
Bazal dendritlər somanın əsasından əmələ gəlir. Bazal dendritik ağac 3-5 ilkin dendritdən ibarətdir. Somadan məsafə artdıqca, bazal dendritlər budaqlanır.[5]
Piramidal hüceyrələr beynin ən böyük neyronlarından biridir. Həm insanlarda, həm də gəmiricilərdə piramidal hüceyrə cisimləri (somalar) orta hesabla 20 mkm uzunluğundadır. Piramidal dendritlərin diametri adətən yarım mkm-dən bir neçə mkm-ə qədər dəyişir. Tək bir dendritin uzunluğu adətən bir neçə yüz mikrometrdir. Budaqlanma səbəbindən piramidal hüceyrənin ümumi dendritik uzunluğu bir neçə sm-ə çata bilər. Piramidal hüceyrənin aksonu çox vaxt daha uzun və geniş şəkildə budaqlanır, ümumi uzunluğu bir çox sm-ə çatır.
Dendritik tikanlar[redaktə | mənbəni redaktə et]
Dendritik tikanlar piramidal hüceyrəyə daxil olan həyəcanverici impulsların çoxunu alır. Dendritik tikanlar ilk dəfə 1888-ci ildə Ramon i Kajal tərəfindən Qolqi üsulu ilə kəşf edilmişdir. Ramón İ Kajal həmçinin neyronal reseptiv səth sahəsinin artması üçün fizioloji rolu təklif edən ilk şəxs idi. Piramidal hüceyrənin səth sahəsi nə qədər böyükdürsə, neyronun böyük həcmdə məlumatı qəbul və inteqrasiya etmək qabiliyyəti bir o qədər yüksəkdir.
Somada dendritik tikanlar yoxdur, ondan uzaqda isə sayı artır.[4] Bir siçovulda tipik apikal dendritdə ən azı 3000 dendritik tikan var.
Orta insan apikal dendritinin uzunluğu siçovulların uzunluğundan təxminən 2 dəfə böyükdür, buna görə də insan apikal dendritində mövcud olan dendritik tikanların sayı 6000-ə qədər ola bilər.[6]
Böyüməsi və inkişafı[redaktə | mənbəni redaktə et]
Differensasiyası[redaktə | mənbəni redaktə et]
Piramidal spesifikasiya beyinin erkən inkişafı zamanı baş verir. Progenitor hüceyrələr subkortikal proliferativ mədəcik zonasında (MZ) və subventrikulyar zonada (SVZ) neyron xəttinə bağlıdır. Yetişməmiş piramidal hüceyrələr kortikal lövhəni tutmaq üçün miqrasiya edir və burada daha da şaxələnir.
Endokannabinoidlər piramidal hüceyrə inkişafına və aksonal yol tapmağa yönəlmiş molekulların bir sinfidir.[7] Ctip2 və Sox5 kimi transkripsiya faktorlarının piramidal neyronların öz aksonlarını yönləndirdiyi istiqamətə töhfə verdiyi göstərilmişdir.[8]
Erkən postnatal inkişaf[redaktə | mənbəni redaktə et]
Siçovulların piramidal hüceyrələrinin doğuşdan sonra həyatda bir çox sürətli dəyişikliklərə məruz qaldığı göstərilmişdir. Doğuşdan sonrakı 3-cü və 21-ci günlər arasında piramidal hüceyrələrin somanın ölçüsündə 2 dəfə, apikal dendritin uzunluğunda 5 dəfə, bazal dendrit uzunluğunda isə 13 dəfə artım müşahidə edilmişdir.
Digər dəyişikliklərə membran potensialının, membran müqavimətinin azalması və fəaliyyət potensialının pik dəyərlərinin artması daxildir.[9]
Siqnalları[redaktə | mənbəni redaktə et]
Əksər digər neyronlardakı dendritlər kimi, dendritlər ümumiyyətlə neyronun giriş sahələridir, akson isə neyronun çıxışıdır. Həm aksonlar, həm də dendritlər çox şaxələnmişdir. Böyük miqdarda budaqlanma neyrona bir çox müxtəlif neyronlara və onlardan siqnal göndərməyə və qəbul etməyə imkan verir.
Piramidal neyronlar, digər neyronlar kimi, çoxlu gərginliyə bağlı ion kanallarına malikdir. Piramidal hüceyrələrdə dendritlərdə çoxlu Na+, Ca2+ və K+ kanalları və somada bir neçə kanal vardır.[10][11] Piramidal hüceyrələrin dendritlərindəki ion kanalları, piramidal hüceyrələrin somasında eyni tipli ion kanallarının xüsusiyyətlərindən fərqli xüsusiyyətlərə malikdir.[12][13] Piramidal hüceyrə dendritlərindəki gərginliyə bağlı Ca2+ kanalları aşağıdakı EPSP-lər və əks fəaliyyət potensialları ilə aktivləşdirilir. Piramidal dendritlər daxilində fəaliyyət potensialının geri yayılma dərəcəsi K+ kanallarından asılıdır. Piramidal hüceyrə dendritlərindəki K+ kanalları fəaliyyət potensiallarının amplitudasına nəzarət mexanizmini təmin edir.[14]
Piramidal neyronların məlumatı inteqrasiya etmək qabiliyyəti onların qəbul etdiyi sinaptik girişlərin sayından və paylanmasından asılıdır. Bir piramidal hüceyrə təxminən 30.000 həyəcanlandırıcı giriş və 1700 inhibitor (IPSP) girişi alır. Həyəcanlandırıcı (EPSP) girişlər yalnız dendritik onurğalarda, inhibitor (IPSP) isə dendritik çubuqlarda, somada və hətta aksonda bitir.
Piramidal neyronlar neyrotransmitter glutamat[5][15] tərəfindən həyəcanlandırıla və GABA (Qamma-aminobutirik turşu) neyrotransmitteri[5]tərəfindən inhibə edilə bilər.
Alt siniflərinin təsnifatı[redaktə | mənbəni redaktə et]
Piramidal neyronlar 400-1000 millisaniyəlik cari impulslara cavablarına görə müxtəlif alt siniflərə bölünür. Bu təsnifatlar RSad, RSna və IB neyronlarıdır.
RSad[redaktə | mənbəni redaktə et]
RSad piramidal neyronlar və ya adi spaykalı adaptiv neyronlar fərdi fəaliyyət potensialı (FP) və sonra hiperpolyarizasiya sonrası potensial tərəfindən işə salınır. Postpotensial müddəti artır, bu da neyronda spayka tezliyi uyğunlaşması (STU) yaradır.[16]
RSna[redaktə | mənbəni redaktə et]
Piramidal neyronlar RSna və ya spaykalı qeyri-adaptiv neyronlar bir impulsdan sonra bir sıra fəaliyyət potensialını işə salır. Bu neyronlarda uyğunlaşma əlamətləri yoxdur.[16]
IB[redaktə | mənbəni redaktə et]
IB piramidal neyronları və ya daxili partlama aktivliyi olan neyronlar, 2-5 sürətli fəaliyyət potensialı ilə Threshold impulslara cavab verir. IB piramidal neyronları uyğunlaşma göstərmir.[16]
Molekulyar təsnifatlar[redaktə | mənbəni redaktə et]
Morfoloji və elektrik piramidal hüceyrələrin xüsusiyyətlərinin tək hüceyrə ardıcıllığı ilə ölçülən gen ifadəsindən çıxarıla biləcəyini göstərən bir neçə tədqiqat var.[17] Bir neçə tədqiqat neyronların müxtəlif xüsusiyyətlərini izah edə bilən gen ifadəsinə əsaslanan siçan [17] və insan [18] neyronlarında tək hüceyrələrin təsnifatını təklif etmişdir. Bu təsnifatlardakı neyron tipləri həyəcanlandırıcı, inhibitor və yüzlərlə müvafiq alt bayta bölünür. Məsələn, insanda 2-3-cü təbəqənin piramidal hüceyrələri FREM3 növü[17] kimi təsnif edilir və çox vaxt HCN kanalı tərəfindən yaradılan yüksək miqdarda Ih cərəyanına[18] malikdir.
Funksiyası[redaktə | mənbəni redaktə et]
Kortikospinal yol[redaktə | mənbəni redaktə et]
Piramidal neyronlar kortikospinal yolun sinir hüceyrələrinin əsas növüdür. Normal motor nəzarəti kortikospinal yolun aksonları ilə onurğa beyni arasında əlaqələrin inkişafından asılıdır. Piramidal hüceyrə aksonları spesifik əlaqələr yaratmaq üçün böyümə faktorları kimi siqnalları izləyir. Düzgün əlaqə ilə, piramidal hüceyrələr görmə ilə idarə olunan motor funksiyasından məsul olan dövrədə iştirak edirlər.[19]
İdrak[redaktə | mənbəni redaktə et]
Prefrontal korteksdəki piramidal neyronlar koqnitiv qabiliyyətlərdə iştirak edir. Məməlilərdə, piramidal hüceyrə mürəkkəbliyi beynin arxa bölgələrindən anterior bölgələrinə qədər artır. Piramidal neyronların mürəkkəblik dərəcəsi, ehtimal ki, müxtəlif antropoid növlərin idrak imkanları ilə əlaqələndirilir. Prefrontal korteksdəki piramidal hüceyrələr ilkin eşitmə qabığı, ilkin somatosensor korteks və ilkin vizual korteksdən gələn məlumatları emal etmək üçün məsuliyyət daşıyırlar, bunların hamısı sensor üsulları emal edir.[20] Bu hüceyrələr korteksin vizual emal sahələrində kompleks obyektlərin tanınmasında da mühüm rol oynaya bilər.[3] Digər növlərlə müqayisədə daha böyük hüceyrə ölçüsü və piramidal neyronların mürəkkəbliyi, hüceyrə təşkili və funksiyasının müəyyən nümunələri ilə birlikdə insan idrakının təkamülü ilə əlaqələndirilir.[21]
Həmçinin bax[redaktə | mənbəni redaktə et]
İstinadlar[redaktə | mənbəni redaktə et]
- ↑ "Arxivlənmiş surət". 2021-02-12 tarixində arxivləşdirilib. İstifadə tarixi: 2023-06-22.
- ↑ Sketchy Group, LLC. "2.3 rhabdovirus". SketchyMedical. 2017-04-13 tarixində orijinalından arxivləşdirilib.
- ↑ 1 2 Elston GN. "Cortex, cognition and the cell: new insights into the pyramidal neuron and prefrontal function". Cerebral Cortex. 13 (11). November 2003: 1124–1138. doi:10.1093/cercor/bhg093. PMID 14576205.
- ↑ 1 2 García-López P, García-Marín V, Freire M. "Three-dimensional reconstruction and quantitative study of a pyramidal cell of a Cajal histological preparation". The Journal of Neuroscience. 26 (44). November 2006: 11249–11252. doi:10.1523/JNEUROSCI.3543-06.2006. PMC 6674523. PMID 17079652.
- ↑ 1 2 3 4 Megías M, Emri Z, Freund TF, Gulyás AI. "Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 pyramidal cells". Neuroscience. 102 (3). 2001: 527–540. doi:10.1016/S0306-4522(00)00496-6. PMID 11226691.
- ↑ Laberge D, Kasevich R. "The apical dendrite theory of consciousness". Neural Networks. 20 (9). November 2007: 1004–1020. doi:10.1016/j.neunet.2007.09.006. PMID 17920812.
- ↑ Mulder J, Aguado T, Keimpema E, Barabás K, Ballester Rosado CJ, Nguyen L, və b. "Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning". Proceedings of the National Academy of Sciences of the United States of America. 105 (25). June 2008: 8760–8765. Bibcode:2008PNAS..105.8760M. doi:10.1073/pnas.0803545105. PMC 2438381. PMID 18562289.
- ↑ Fishell G, Hanashima C. "Pyramidal neurons grow up and change their mind". Neuron. 57 (3). February 2008: 333–338. doi:10.1016/j.neuron.2008.01.018. PMID 18255026.
- ↑ Zhang ZW. "Maturation of layer V pyramidal neurons in the rat prefrontal cortex: intrinsic properties and synaptic function". Journal of Neurophysiology. 91 (3). March 2004: 1171–1182. doi:10.1152/jn.00855.2003. PMID 14602839.
- ↑ Spruston N. "Pyramidal neurons: dendritic structure and synaptic integration". Nature Reviews. Neuroscience. 9 (3). March 2008: 206–221. doi:10.1038/nrn2286. PMID 18270515.
- ↑ Georgiev DD, Kolev SK, Cohen E, Glazebrook JF. "Computational capacity of pyramidal neurons in the cerebral cortex". Brain Research. 1748. December 2020: 147069. doi:10.1016/j.brainres.2020.147069. PMID 32858030 (#bad_pmid).
- ↑ Golding NL, Mickus TJ, Katz Y, Kath WL, Spruston N. "Factors mediating powerful voltage attenuation along CA1 pyramidal neuron dendrites". The Journal of Physiology. 568 (Pt 1). October 2005: 69–82. doi:10.1113/jphysiol.2005.086793. PMC 1474764. PMID 16002454.
- ↑ Remy S, Beck H, Yaari Y. "Plasticity of voltage-gated ion channels in pyramidal cell dendrites". Current Opinion in Neurobiology. 20 (4). August 2010: 503–509. doi:10.1016/j.conb.2010.06.006. PMID 20691582.
- ↑ Magee J, Hoffman D, Colbert C, Johnston D. "Electrical and calcium signaling in dendrites of hippocampal pyramidal neurons". Annual Review of Physiology. 60 (1). 1998: 327–346. doi:10.1146/annurev.physiol.60.1.327. PMID 9558467.
- ↑ Wong, R. K. S.; Traub, R. D., NETWORKS | Cellular Properties and Synaptic Connectivity of CA3 Pyramidal Cells: Mechanisms for Epileptic Synchronization and Epileptogenesis // Schwartzkroin, Philip A. (redaktor), Encyclopedia of Basic Epilepsy Research (ingilis), Oxford: Academic Press, 2009-01-01, 815–819, doi:10.1016/b978-012373961-2.00215-0, ISBN 978-0-12-373961-2, İstifadə tarixi: 2020-11-18
- ↑ 1 2 3 Franceschetti S, Sancini G, Panzica F, Radici C, Avanzini G. "Postnatal differentiation of firing properties and morphological characteristics in layer V pyramidal neurons of the sensorimotor cortex". Neuroscience. 83 (4). April 1998: 1013–1024. doi:10.1016/S0306-4522(97)00463-6. PMID 9502243.
- ↑ 1 2 Berg J, Sorensen SA, Ting JT, Miller JA, Chartrand T, Buchin A, və b. "Human neocortical expansion involves glutamatergic neuron diversification". Nature. 598 (7879). October 2021: 151–158. doi:10.1038/s41586-021-03813-8. PMC 8494638 (#bad_pmc). PMID 34616067 (#bad_pmid).
- ↑ Kalmbach BE, Buchin A, Long B, Close J, Nandi A, Miller JA, və b. "h-Channels Contribute to Divergent Intrinsic Membrane Properties of Supragranular Pyramidal Neurons in Human versus Mouse Cerebral Cortex". Neuron. 100 (5). December 2018: 1194–1208.e5. doi:10.1016/j.neuron.2018.10.012. PMC 6447369. PMID 30392798.
- ↑ Salimi I, Friel KM, Martin JH. "Pyramidal tract stimulation restores normal corticospinal tract connections and visuomotor skill after early postnatal motor cortex activity blockade". The Journal of Neuroscience. 28 (29). July 2008: 7426–7434. doi:10.1523/JNEUROSCI.1078-08.2008. PMC 2567132. PMID 18632946.
- ↑ Baker A, Kalmbach B, Morishima M, Kim J, Juavinett A, Li N, Dembrow N. "Specialized Subpopulations of Deep-Layer Pyramidal Neurons in the Neocortex: Bridging Cellular Properties to Functional Consequences". The Journal of Neuroscience. 38 (24). June 2018: 5441–5455. doi:10.1523/JNEUROSCI.0150-18.2018. PMID 29798890.
- ↑ Galakhova AA, Hunt S, Wilbers R, Heyer DB, de Kock CP, Mansvelder HD, Goriounova NA. "Evolution of cortical neurons supporting human cognition". Trends in Cognitive Sciences (English). 26 (11). November 2022: 909–922. doi:10.1016/j.tics.2022.08.012. PMC 9561064 (#bad_pmc). PMID 36117080 (#bad_pmid).
{kind=link}
{kind=link}